Structure of HIV outer shell determined
31 Jan 2011
The structure of the protein package that delivers the genetic material of the human immunodeficiency virus (HIV) to human cells has been mapped by the The Scripps Research Institute and the University of Virginia.
The work is the culmination of studies carried out over the last decade looking at different portions of the capsid — the cone-shaped container, of the virus.
The final piece of the puzzle, described in an article published in the journal Nature on January 20, 2011, details the structure of the two ends of the cone.
“This paper is a real milestone for research from our group,” says the study’s senior author Mark Yeager, M.D., Ph.D., a Scripps Research professor and staff cardiologist and chair of the Molecular Physiology and Biological Physics Department at The University of Virginia School of Medicine.
A detailed description of the complete HIV capsid will provide a roadmap for developing drugs that can disrupt its formation and thus prevent infection by HIV.
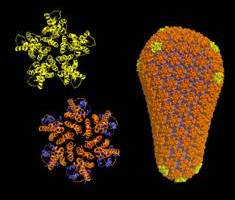
| The new detailed description of the
complete HIV capsid. The capsid of HIV is a conical fullerene shell that comprises about 250 hexamers (orange) and exactly 12 pentamers (gold) of the viral CA protein. In the new study, the researchers determined X-ray crystal structures of these building blocks, which enabled modeling of the complete capsid at atomic resolution. The continuously varying lattice curvature in the fullerene cone can be explained simply by two rigid body rotations around two assembly interfaces of CA. (Graphics by Owen Pornillos, Barbie Ganser-Pornillos, Kelly Dryden, and Mark Yeager.) |
Assembling the package
HIV binds to receptors on human cells and then delivers the capsid inside them. Once inside a cell, the capsid comes apart, releasing its precious cargo — the virus’s genetic material.
HIV then sabotages the cell machinery to make many copies of its genes and proteins. As new viruses are made, the genetic material is packaged into spherical immature capsids that HIV uses to escape from the infected cell. But before these newly released viruses can infect other cells, the immature capsid undergoes a dramatic rearrangement to form the mature, cone-shaped shell.
If formation of the mature capsid is disrupted, the virus is no longer infectious. Thus, new drugs targeting capsid formation could provide valuable additions to the arsenal of existing drugs against HIV.
A "floppy" bridge
To develop drugs that disrupt capsid formation, however, scientists first need to know precisely how it is formed. One technology researchers use to obtain detailed structures of biological molecules is X-ray crystallography. This technique requires growing crystals of a molecule and then bombarding the crystals with X-rays to determine the positions of all the atoms.
But unlike the cone-shaped capsids of other viruses, such as the poliovirus, which have a rigid, symmetrical structure that obediently assembles into crystals, the HIV capsid is flexible and can adopt slightly different shapes.
Part of the reason for this flexibility is the protein that makes up the HIV capsid, the CA protein, consists of two ends held together by a “floppy” bridge.
In the capsid, each CA protein joins hands with other CA proteins, forming groups of five or six proteins. The main body of the capsid contains about 250 of the six-fold units or hexamers. Each end of the cone is then closed off by either five or seven smaller five-fold units or pentamers.
“It is impossible to grow crystals of the entire HIV capsid,” says Yeager. As a result, his team used a “divide and conquer approach.”
Divide and conquer
Working with husband-and-wife team Owen Pornillos and Barbie Ganser-Pornillos, investigators in his lab, Yeager partitioned the HIV capsid into smaller components, then determined their respective structures.
Yeager's group started by focusing on the structure of the CA hexamer. A breakthrough came in a 2007, when the group viewed the CA hexamers with a powerful electron microscope. Guided by information from that structure, in 2009 the team managed to trick the CA hexamers into forming crystals. The researchers were then able to determine the particles’ structures at 2-Angstrom resolution (one Angstrom equals one ten-billionth of a meter).
Having cracked the atomic structure of the hexamer, the investigators turned their attention to the more elusive pentamers.
Structure of the Pentamer
In this latest study, Yeager, Pornillos, and Ganser-Pornillos used techniques similar to those they had applied to the hexamers to obtain the crystal structures of the CA pentamers.
The new structure reveals that five CA proteins link hands at one end, called the N-terminal domain (NTD), to form a circle. The opposite ends of the CA proteins, called C-terminal domain (CTD), form a floppy belt around this central core. Then, CTD links to CTD to connect adjacent pentamers.
The structure reveals flexibility and mobility both between the central core and belt within each pentamer and at the CTD-CTD interfaces of adjacent pentamers. The CTD subunits can rotate relative NTDs. “As a result, each ring can adopt slightly different angles relative to its adjacent rings,” says Pornillos, first author of the paper.
The structure of the pentamers is remarkably similar to that of the hexamers, notes Pornillos, with one important difference. Because pentamers are smaller than hexamers, the amino acids, the building blocks of proteins, at the center of the pentamer ring are closer together than in the hexamer.
Many amino acids have positive or negative charges. When two amino acids with the same charge are close together they tend to push each other away. One amino acid in the CA protein, called arginine, with a positive charge, sits smack in the middle of both the hexamer and pentamer ring.
Because in the pentamer the arginines are packed much closer together, they repel one another, making the pentamer a less stable structure than the hexamer. This may explain why there are many more hexamers in the mature HIV capsid compared to pentamers.
The only place where pentamers are likely to form is at the capsids’ ends, where the linked CA proteins have to bend dramatically to close off the capsid—a feat the pentamer is more apt to perform.
“Arginine is the critical switch between hexamer and pentamer formation,” says Yeager. “We can finally explain why the CA protein would make one or the other.”
An atomic model of the HIV Capsid
Having solved the atomic structures of both CA hexamers and pentamers, Yeager and colleagues for the first time were able to build a complete atomic model of the mature HIV capsid.
The researchers now plan to further refine the model using sophisticated computer programs to determine the stability of the structure in different regions and to identify possible “weak” points they can target using newly designed drugs.
They will also begin studying the structure of the immature capsid to determine how this version of the capsid transitions to the mature form — a step in the virus lifecycle that has remained mysterious.
“We don’t have the full story yet, but we have volume one,” says Yeager.
Reference
Owen Pornillos, Barbie K. Ganser-Pornillos, Mark Yeager. Atomic level modeling of the HIV capsid. Nature, January 20, 2011.